2018/04/09
酵母が冷凍保存後も高い発酵力を維持する仕組みを解明
変性タンパク質を分解する酵素が重要な役割
~冷凍耐性や発酵力が向上したパン酵母の育種に期待~
【概要】
奈良先端科学技術大学院大学(学長:横矢直和)先端科学技術研究科バイオサイエンス領域ストレス微生物科学研究室の高木博史教授のグループは、テーブルマーク株式会社(代表取締役社長:川股篤博)との共同研究により、パン酵母が冷凍保存後も高い発酵力を発揮するためには、細胞内で冷凍により傷害を受け変性したタンパク質を分解するという役割を持つ酵素複合体が重要であることを明らかにしました。
酵母※1は発酵産業において広く利用されている微生物ですが、製パンに用いる酵母(パン酵母)は種々の製パンの過程で冷凍、高ショ糖、高温乾燥などのストレス環境にさらされながら、高い発酵力(炭酸ガス発生量で評価)を維持しています。したがって、パン酵母が発酵力を保持したまま冷凍状態で流通することができれば、製パン産業において有用な技術になると期待されています。
本研究では、まずパン酵母の冷凍保存が、発酵中の細胞の遺伝子発現に及ぼす影響を調べました。その結果、冷凍保存後の発酵力が低下した株では、細胞内でタンパク質分解を行う巨大な酵素複合体(プロテアソーム※2)に関連する遺伝子のほとんどで発現が減り、ストレスにより変性したタンパク質が分解されずに蓄積していました。このことから、冷凍保存によりプロテアソームの機能が低下していることが分かりました。次に、プロテアソームの機能低下を引き起こす原因を解析したところ、冷凍保存後に発酵力が低下した株では、プロテアソーム関連遺伝子の発現を引き起こす転写因子であるPdr3タンパク質※3にアミノ酸置換を伴う変異が入り、Pdr3タンパク質の標的になる遺伝子の発現も顕著に減っていました。また、このPdr3タンパク質変異体を実験室酵母で発現させたところ、冷凍保存後の発酵力や生存率が有意に下がりました。
以上の結果から、冷凍保存によるパン酵母の発酵力低下はプロテアソームの機能欠損が原因であることが明らかになりました。また、パン酵母が冷凍ストレスを克服し、高い発酵力を維持するためには、プロテアソームの機能が重要な役割を果たしており、この機能を強化することでパン酵母の冷凍耐性や発酵力が向上する可能性が示されました。
本研究で得られた知見は、プロテアソームの機能強化による「冷凍ストレス耐性パン酵母」の育種技術への応用や、細胞が冷凍ストレスに応答し、生命機能を保持するメカニズムの理解に繋がる有用なものであると考えられます。また、本研究の成果はパン酵母をはじめとする産業酵母について、タンパク質を変性させる様々なストレスに対する耐性や発酵力の向上に応用できる可能性があります。
この研究成果は、米国微生物学会の学会誌であるApplied and Environmental Microbiology誌オンラインサイトに平成30年4月6日付けで掲載されました。
【解説】
研究背景
パン酵母は約1 兆4,000 億円もの製パン産業を支える微生物であり、年間で200 万トンほど製造されています。多様な製パン法に対応可能なパン酵母の開発は製パン業界における重要な技術です。例 えば、オーブンフレッシュベーカリーやコンビニエンスストアでは、製パン過程の途中で生地を冷凍する方法で製造されるパンが販売されています。また、菓子パンの生地には小麦粉重量あたり30-40% ものショ糖が含まれています。さらに、細胞内の水分含量を下げて製造するドライイーストは長期間の貯蔵や輸送経費の削減を可能にします。このように、パン生地の発酵やパン酵母の製造過程で、酵母の細胞は冷凍、高ショ糖、高温乾燥などのストレス環境にさらされています。
このようなストレス下では、タンパク質など生体高分子の構造や機能が失われ、パン酵母の有用機能(炭酸ガスの発生、味・風味成分の生成など)が制限されます。したがって、パン酵母に高度なス トレス耐性を付与することにより、長期保存可能な冷凍生地や菓子パン生地に適した「冷凍耐性イースト」「高糖耐性イースト」、耐久性の強い「ドライイースト」の開発が可能になります。また、パン 酵母が発酵力を保持したまま冷凍状態で流通することができれば、製パン産業において有用な技術になると期待されています。そこで本研究では、冷凍ストレスがパン酵母の発酵力と遺伝子発現に及ぼす影響を明らかにし、冷凍保存後の発酵力に関与する遺伝子の同定と機能解析を行いました。
研究結果
テーブルマーク社のパン酵母二倍体株(45 株およびYF 株)を2 週間冷凍保存した後、高ショ糖生地を模倣した液体培地で発酵速度を測定したところ、YF 株は45 株よりも顕著に発酵力が低下していました。そこで、このYF 株に特異的な遺伝子発現の特徴を解析した結果、プロテアソームを介したタンパク質分解に関連する遺伝子の発現がYF 株において顕著に低下していることを見出しました。プロテアソームはストレスにより傷害を受け、変性したタンパク質を効率良く除去するために必要なメカニズムであることから、YF 株では冷凍保存により傷害を受けたタンパク質が蓄積し、発酵力が低下している可能性が示されました。この仮説を検証するために、45 株とYF 株における変性タンパク質の蓄積に関する解析を行ったところ、YF 株では45 株と比較して熱ショックや冷凍により変性するタンパク質が多く蓄積していることが明らかになりました(図1)。
次に、次世代シーケンサーを用いてYF 株のゲノム解析を行い、プロテアソームの機能低下を引き起こす原因遺伝子を探索しました。その結果、YF 株ではプロテアソーム関連遺伝子の転写活性化因子として知られるPdr3 タンパク質の遺伝子(二倍体なので2 個存在)の片一方にアミノ酸置換を伴う変異が生じており(Ala148Thr/Ala229Val/His336Arg/Leu541Pro)(図2)、Pdr3 タンパク質の標的遺伝子(RPN4, PDR5, SNQ2)の発現量も顕著に低下していました。さらに、このPdr3 タンパク質変異体(Ala148Thr/Ala229Val/His336Arg/Leu541Pro)を実験室酵母で発現させたところ、YF 株と同様に冷凍保存後の発酵力や生存率が有意に低下しました(図3)。また、YF 株におけるPdr3 タンパク質遺伝子の変異はドミナントネガティブ※4型であることも示唆されました。
以上の結果から、Pdr3 タンパク質の機能欠損がYF 株におけるプロテアソームの機能低下と冷凍保存後の発酵力低下の原因であることが証明されました。また、冷凍ストレス応答におけるプロテアソームの機能の重要性が明らかになり、プロテアソームの機能強化によって冷凍ストレス耐性酵母の育種への可能性を提唱することができました。
研究意義
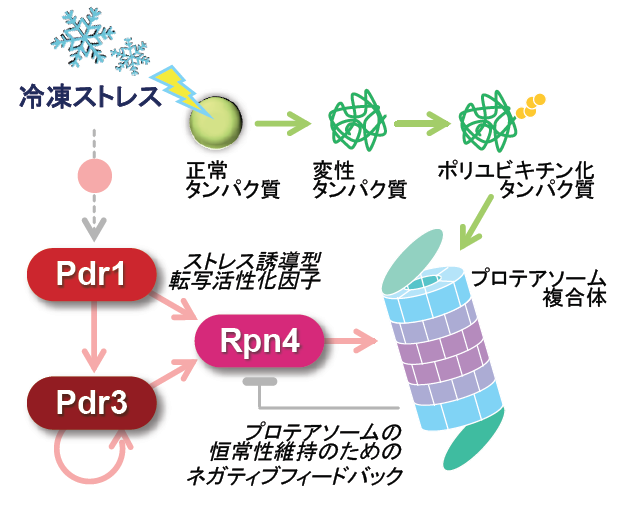
以上の結果から、冷凍保存によるパン酵母の発酵力低下はプロテアソームの機能欠損が原因であること、およびパン酵母が冷凍ストレスを克服し、高い発酵力を維持するためには、プロテアソームの機能強化が重要であることを明らかにしました。今回の研究成果に基づき、酵母における冷凍ストレス応答メカニズムの新しいモデルが考えられます(図4)。その鍵を握るのは転写活性化因子(Pdr1タンパク質、Pdr3 タンパク質、Rpn4 タンパク質※5)を介したプロテアソームの発現誘導であり、YF株はこのうちPdr3 タンパク質の遺伝子にドミナントネガティブ型の変異を持つことで冷凍ストレス耐性を獲得できなくなったと思われます。今回の研究で得られた知見は、プロテアソームの機能強化による「冷凍ストレス耐性パン酵母」の育種技術への応用や、細胞が冷凍ストレスに応答し、生命機能を保持するメカニズムの理解につながる有用なものであると考えられます。また、本研究の成果はパン酵母をはじめとする産業酵母について、タンパク質を変性させる様々なストレスに対する耐性や発酵力の向上に応用できる可能性があります。
さらに、酵母のストレス耐性機構に関する知見と技術は、様々な発酵・醸造食品(パン類、酒類、醤油、味噌、酵母エキスなど)の製造にとどまらず、バイオエタノールを含む主要な酵母利用産業の高度化・効率化に役立つ技術への展開も期待されます。
【掲載論文】
タイトル:Importance of Proteasome Gene Expression during Model Dough Fermentation after Freezing Preservation of Baker's Yeast Cells
(和訳:パン酵母の冷凍保存後の発酵力に重要なプロテアソーム関連遺伝子の発現)
著者:Daisuke Watanabe1), Hiroshi Sekiguchi2), Yukiko Sugimoto1), Atsushi Nagasawa2), Naotaka Kida2),Hiroshi Takagi1)
所属:1)奈良先端科学技術大学院大学、2)テーブルマーク株式会社
掲載誌:Applied and Environmental Microbiology
naister:http://library.naist.jp/dspace/handle/10061/12548(NAIST Academic Repository:naistar)
【用語解説】
※1:酵母
酵母のうち基礎研究のモデル株を実験室酵母と称する。実験室酵母では遺伝解析やゲノム解析による知見が蓄積され、遺伝子操作技術も確立しているが、発酵力は弱く発酵食品などの生産には不適である。一方、製パン・醸造、バイオエタノール生産などに用いられる菌株を産業酵母と称する。産業酵母は発酵性や生育速度の優れた株が選択されているが、遺伝特性や倍数性が異なり、実験室酵母の知見や技術が活用されていない。本研究では、産業酵母のパン酵母を使用した。
※2:プロテアソーム
真核生物の細胞質および核内に存在し、タンパク質を分解する巨大な酵素複合体である。多くのサブユニットから構成され、細胞内で不要になったり、変性したタンパク質にユビキチンという小さなタンパク質が付加すると、これが目印になり選択的な分解と再利用を行う。多くの場合、反応にATPを必要とし、細胞内の基質もいくつか明らかにされている。
※3:Pdr3タンパク質
元々は部分的に相同性を示すPdr1タンパク質とともに、酵母の多剤耐性(Pleiotropic drug resistance)に関与する転写因子として同定され、薬剤の排出に必要なABCトランスポーター(Pdr5タンパク質、Snq2タンパク質など)をコードする遺伝子の発現を誘導することが知られていた。その後、薬剤耐性の獲得以外にも、ストレス応答などの幅広い役割を有することが明らかになっている。
※4:ドミナントネガティブ
遺伝子に変異が入った際に、変異型の遺伝子産物(タンパク質など)が正常型の遺伝子産物よりも量が多い、または正常型を阻害する働きがあるなどの理由で、変異型の遺伝子産物の働きが優性になること。特に、変異型と正常型の遺伝子産物が複合体を形成する場合に起こりうる。
※5:Rpn4タンパク質
プロテアソーム関連遺伝子の発現において中心的役割を果たす転写因子。Rpn4の発現は熱ショックや酸化ストレス、アミノ酸アナログなどのタンパク質毒性ストレス、DNA損傷により誘導され、プロテアソーム量を増加させる。また、Rpn4自身がプロテアソームの基質であり、プロテアソーム量が増加するとRpn4量が減少するネガティブフィードバックが存在する。
【本研究内容についてコメント出来る方】
喜田 直孝[テーブルマーク株式会社 食品開発センター 研究開発部長]
TEL: 03-5705-7554 FAX: 03-5705-8775 E-mail: [email protected]
【本プレスリリースに関するお問い合わせ先】
奈良先端科学技術大学院大学 先端科学技術研究科
バイオサイエンス領域 ストレス微生物科学研究室 教授 高木 博史
TEL: 0743-72-5420 FAX: 0743-72-5429 E-mail: [email protected]
【解説図】

左:熱ショック(42℃, 15分)後、右:冷凍(2週間)後
ユビキチン抗体で変性タンパク質を検出

(DBD:DNA結合ドメイン、ID:阻害ドメイン、AD:活性化ドメイン)
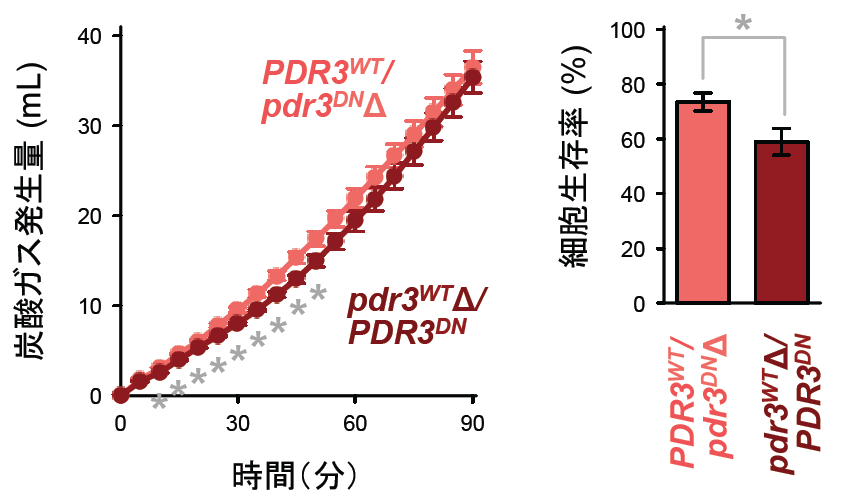
PDR3WT/pdr3DNΔ:野生型Pdr3のみ発現、pdr3WTΔ/PDR3DN:変異型Pdr3のみ発現
左:冷凍(2週間)後の発酵力、右:冷凍(2週間)後の生存率
*印は有意差があることを示す